Collaborative Proposal Development Towards Sustainable Communities
 James Furze
James Furze
University of the West of England, Faculty of Environment and Technology
James.Furze@uwe.ac.uk
.
 Quanmin Zhu
Quanmin Zhu
Professor at University of the West of England, Faculty of Environment and Technology
Quan.Zhu@uwe.ac.uk
.

Feng Qiao
ShenyangJianzhu University, Faculty of Information and Control Engineering
.
[soundcloud url=”https://api.soundcloud.com/tracks/118106567″ params=”color=ff6600&auto_play=false&show_artwork=true” width=”100%” height=”166″ iframe=”true” /]
Abstract: Sustainable communities require consideration from different perspectives. Maintaining a sustainable community using a systematic approach enables the meeting of multiple objectives benefitting ecological systems and human populations, who are dependent on the stability of them. We list examples of interdisciplinary research, benefitting collaborative proposal formation and generating and sustaining wide audiences within research and in communities. Mathematic integrated algorithm construction details qualitative characteristics of plant species integration into common models and quantification to dictate a control strategy based approach of relevance to over 300 000 plant species. Field-data simulations formed use genetic programming techniques and functional mathematic approximation via Gaussian process models. Implemented algorithms were made use of as the modeling basis on which we developed a geographic information system (GIS), summarizing plant distribution using principal variables of the water-energy dynamic. The primary metabolic process of plants was ordered within each of the 7 plant strategy environments. Dominance of 3 carbon photosynthesis was seen in ruderal through stress tolerant-ruderal to competitive-ruderal environments where water is the key part of the water-energy dynamic in effect. Dominance of 4 carbon photosynthesis was found in competitive and competitive-stress tolerant-ruderal environments. Dominance of crassulacean acid metabolism was found in competitive-stress tolerant and stress tolerant environments where energy is the key part of the water-energy dynamic in effect. Algorithms were tested by varying the dynamic variables inputted into the models, in which case the refined mathematic technique resulted in an incorrect summation. In conclusion control strategy offered a viable approach to describe plant characteristics using concise statements. It assisted in development of a modeled GIS to use as a tool for policy makers. The algorithmic basis of the GIS has great potential in simulation development. Linking of different time scenario climatic data, topographic data, image processing, map processing, species occurrence records and finally digital mapping enhance novel software formulation for design of control systems with both mathematical and geographical global inferences. Collaborative proposals between international research departments in control science, geography, operational research, plant science, computing / machine vision, co-operative systems and environmental planning law have potential subject unification, providing strong recommendations of policy formation with socio-economic connotations. These areas are key to motivating multi-participation in future research, which ensures natural and human sustainable communities in different settings.
Keywords: Sustainable communities, systematic approach, ecological systems, algorithm, plant species, programming, functional approximation, photosynthesis, strategy, collaborative proposals, policy makers.
.
Introduction
In ecological terms, sustainable communities are those that are made up of many parts the growth of which develop with a synergistic function to benefit the future population growth of the groups of organisms found within. The abiotic and biotic conditions of the community share relationships with the components of the community at various levels (including the ecosystem, habitat, groups of individuals and individual species). Thus numbers and types of individual species may be used as an indicator of the conditions in which a community is found. These relationships are seen to follow both linear and non-linear relationships. Arrhenius (1921) demonstrated the species-area relationship where the number of species and the area of a location (with exponents of environmental conditions and taxa types) share a linear relationship. Arrhenius was followed by authors who applied a species-area relationship and defined specific terms to different island or mainland areas (MacArthur and Wilson, [ch.1] 1967; Simberloff, 1974). It is been accepted that the nonlinear, discrete relationships of species patterns are multi-factorial, the most important factors include genetic pressures from within the species population / other species, spatial restrictions and resources required for growth. As ecosystems are made up of many layers of organization in a hierarchical structure, it is logical to assume that the primary producing level of the ecosystem is fundamental to the number of species found in subsequent trophic levels.
In order to clarify essential elements required for growth within plant communities, modeling has been applied. Biological, chemical and geographical areas have been integrated with use of engineering based technique. Plant strategies, plant metabolism and plant life-form type have all been characterized and stochastic patterns have been distributed on a global scale (Furze, Zhu, Hill and Qiao, 2013a; Hawkins et al., 2003; Schölzel and Friedrichs, 2008). It is envisaged that the latter refined mathematic technique enables greater understanding of trophic levels, of the conditions in which they exist and of subjects being processed in order to reach the optimal decision on which multi-objective optimization of plant species characteristics is based.
Leading experts in the field of plant species characterization cross boundaries of the contributing subjects and combine disciplines in order to strengthen their arguments and generate novel subject areas for future research (Prusinkiewicz and Lindenmayer, 1990; Grime and Pierce, 2012; Furze, Zhu, Qiao and Hill, 2013b). Areas of importance include specific morphology development, plant strategy and community functional organization.
Categorization and mapping of global locations in terms of primary producing species is of great importance. Further, existing and predictive modelling of species distributes are an increasingly important established method of informing policy recommendations of ecosystem and conservation significance (Trauth, [ch. 1], 2006). However a key element for policy makers is the question of how we may draft and further implement the policies themselves. Collaborative proposals of multiple research areas with communities and governmental bodies provide an opportunity for engagement of individuals with research and the setting of goals which safeguard natural population diversity and our place within it.
This paper proposes a collaborative program with sustainability and enhancement of conservation status at its core. The authors would like to highlight that there are many predictive functions offered with use of mathematic modeling. These enable foresight within populations for natural species organization application with variable pressures being imposed. There are additionally benefits of optimal plant populations meeting the requirements (e.g. food, medicinal, and material) in human populated areas.
The aim of this paper is to describe areas with synergistic functions for progress within interdisciplinary areas of plant characterization and to explore predictive benefits for natural communities and human awareness. Objectives of this collaborative proposal are to unify plant science, biogeography, mathematics and operational research; to make formal statements of plant distribution in terms of global distribution patterns of plant strategies, life-forms and metabolic pathways and ascertain whether such statements can be justified and made with use of concise modeling frameworks / tools.
It is hoped that funding in contributing disciplines and from national governing bodies will be generated to facilitate the program set out in the following section. The remainder of this paper is structured as follows: the methods section presents the key parts of the proposal. The results section shows global distribution of plant strategies. The discussion section details the result and gives other potential applications. Finally future research and its potential for human sustainability proposals concludes the paper.
Method
Mathematic integrated algorithm construction
Following review of mathematical methods for plant characterization (Furze et al., 2013b) we choose to integrate characteristics of plant species of plant strategies (Grime, Hodgson and Hunt, [ch.1] 1995), plant life-form (Raunkier, 1934) and photosynthesis (Salisbury and Ross, [ch. 12], 1992) using a Takagi-Sugeno-Kang modeling framework (Takagi and Sugeno, 1985).
We constructed algorithms for plant distribution on a global scale, using field-based data of individual plant occurrences for consequent numbers from the publicly stored multiple-interface portal of the Global Biodiversity Information Facility (www.gbif.org, accessed December 2010). After consideration of a range of elements of the water-energy dynamic (Hawkins et al., 2003), we made use of quarterly climatic data of 1961-90, mean annual precipitation and mean temperature (New et al., 1999). Altitude data of GTOPO30 was also included (www.usgs.gov, accessed January 2012) to accord for the variable effect of water and energy at different heights (Wright, 1983; Bhatterai and Vetaas, 2003). We summarized distribution of plant species in the richest locations (Barthlott, Mutke, Rafiqpoor, Kier and Kreft, 2005) with relevance to over 300 000 species and presented algorithms for characteristics of plant-strategy, plant photosynthetic type and plant life-form (Furze, Zhu, Qiao and Hill, 2012; Furze, Zhu, Qiao and Hill, 2013c; Furze, Zhu, Qiao and Hill, 2013d). The algorithms were tested by viewing 2 dimensional and 3 dimensional surfaces of the algorithms. Varying 1 or more of the driving dynamic factors resulted in either a static response or the too high/too low a result being achieved for the plant characteristic. A summary of plant species strategy balance within each environment is given in Table I. There are three main types of photosynthesis. Ordering of these three types is given within each strategy based environment. Raunkier (1934) plant life-forms were also extrapolated throughout the 7 environments, these plant characteristics show continually expressed distribution patterns which were summarized by imposing the boundaries of Rastrigin’s function, Gaussian process model to give spectra, for the results of this standardized analysis see Furze et al., 2013d.
Geographic information system build
We constructed a geographic information system (GIS) using our algorithmic foundation detailed above. The technical computing platform Matlab (Version 2010a ©) mapping function was used as the framework for the GIS, enabling a plant characteristic based global map to be presented. Each location was selected by selecting the tile code (e.g. W100N40 for Ecuador), number of columns from left to right and row number from top to bottom. Resolution of the areas may be enhanced to include the latitude, longitude of each area with use of Matlab (Version 2010a ©) code, for details contact the first author. The summary GIS tool is presented in Figure I.
Plant science-mathematic expansion
We expanded the primary numbers of plant characteristics (e.g. 7 plant strategies expanded to 20 rudiments) following a genetic programmed dispersal of the rudimentary characters. Hence we obtained first and second order equations for plant characteristics, following T-S-K modeling and multi-objective genetic algorithms respectively. We made a concise summary of the expansion process in construction of a hybrid multi-objective genetic algorithm (Furze et al., 2013a). Construction of a Pareto front of the combined objective distribution of plant strategies enabled linear and quadratic expressions of utopia to be formed. The significance of the latter is that a predictive function of climatic systems versus species numbers was enabled. Further exploration of combined objective planes were enabled with use of a process functional approximation of the distribution of plant life-forms (Büche, Schraudolph and Koumoutsakos, 2005; Furze et al., 2013d). The use of this final step was key for future research and is covered in the discussion section.
Results
After proposing the sum of strategy types equates to 1 in order to classify an environment of plant species (Furze et al., 2013b), we were able to produce the generalized approximation of the proportions of plant strategy types present in each of the 7 plant strategy environments (Furze, Zhu, Qiao and Hill, 2013c).
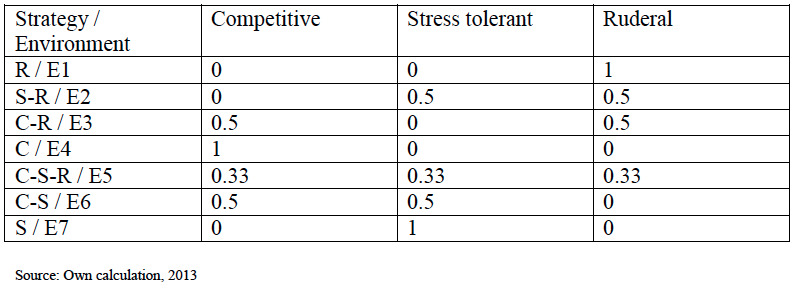
Table I Plant species competitive-stress tolerant-ruderal balance in numerical form
R refers to Ruderal, S-R refers to stress tolerant-ruderal, C-R refers to competitive-ruderal, C refers to competitive, C-S refers to competitive-stress tolerant, S refers to stress tolerant. Example environments are (E1) Ecuador, South America, (E2) Guyana, (E3) Cuba, (E4) Democratic Republic of the Congo, Africa, (E5) Georgia, (E6) Azerbaijan, Europe, (E7) Macedonia (Europe). Justification of the use of these example areas and their consequent individual occurrence numbers has been stated in recent publications (Furze et al.,2013c; Furze, Zhu and Hill, 2013).
In reality, the existence of a pure strategy without any elements of the others very rarely exists due to the polyploidy level of plant species. By extension to an ecosystem approach this means that in plant-strategy environments it is likely that levels of competitive and of stress tolerant species do exist in ruderal environments. The limit after which the strategy element is detected is 1/3.
Figure I Summary GIS tool for policy makers

Environments E1-E7 were indicated on the above GIS, representing dominant strategies in the locations to form the environments. Each environment is highlighted with a different colour in order to reflect the combination of water and energy, which prevail in the location. Colours are ordered from blue (E1), grey blue (E2), light blue (E3), green (E4), rose (E5, orange (E6) and red (E7).
The T-S-K algorithms can be broken down into rules of the water-energy and altitude variables for the above areas, which were given in Furze et al., 2013c.
Processing of photosynthetic types with the environments E1-E7 indicates that according to the water-energy dynamic effect on photosynthesis, photosynthesis is ordered in the following way:
C3 ≥ C4 ≥ CAM for all E1; E2; E3 (1)
C4 ≥ C3 ≥ CAM for all E4; E5 (2)
CAM ≥ C4 ≥ C3 for all E6; E7 (3)
Where ≥ is greater than or equal to, C3 is 3 carbon photosynthesis, C4 is 4 carbon photosynthesis, CAM is crassulacean acid metabolism and E1-E7 are defined as in Table I.
Conclusions
Greater numbers of plant species in ruderal based environments were found in equatorial areas where the level of water (represented by mean annual precipitation) related variables are high, whereas competitive and stress tolerant based plant environments were found in locations where energy (represented by mean annual temperature) are expressed with greater weight acting on the distribution. The areas of areas of competition and stress tolerance were principally ordinated above the equator, e.g. Macedonia. The finite distribution of plant species requires further research in areas of physical and biochemical characterization. It is suggested that Gaussian patterns identified may be further extrapolated into discrete distributes as stochastic, discrete functions enable characterization of plant species in terms of different secondary metabolic groups (e.g. by plant product). It is suggested that use of machine vision technique of viewing colour spectra of climatic images, is to be of use in establishing knowledge bases for algorithmic construction towards metabolic characterization at increased resolution within broadly characterized areas.
The algorithmic basis of GIS shows great potential in simulation development as algorithms are constructed from initial knowledge bases, which are publicly available with use of the internet. Linking of different time scenario climatic data, topographic data, image processing, map processing, species occurrence records and finally digital mapping are techniques by which new software may be formulated for design of control systems with both mathematical and geographical global inferences. This represents novel simulation development using driving dynamic factors of the water-energy dynamic and continuously monitored numbers of plant species occurrence. The inferences of such a GIS allow the unification of multiple subject areas to substantiate policy formation and enable subject development in the component areas (e.g. geography, biochemistry, physiology and mathematics). Generally GIS offer cost and time savings involved in data collection and interpretation, may lead to better decision-making and improved communication.
Collaborative proposals between research departments offering expertise in control science, geography, operational research, plant science, computing / machine vision, co-operative systems and environmental planning law have great potential in that they unify subjects to provide strong recommendations of policy formation with socio-economic connotations. There are also many roles that the general public can become involved in such research, including recording and storing species presence data, surveying their living needs for a requirement of certain plant products (e.g. medicinal, food or structural materials). Implementation of the algorithmic approach to provide functional approximation of plant metabolites will provide highly informative material for policy makers (Furze et al., 2013d).
Further benefits of this work are the advancement of research in multiple subject areas and improvement of knowledge bases used. The work contributes to safeguarding of indigenous and natural populations reliant on plant species for their living requirements. Beneficial effects towards long-term sustainability in ecosystems trophic levels reliant on rich plant distribution are felt (Kreft and Jetz, 2007; Jenkins, Pimm and Joppa, 2013; Kraft and Ackerly, 2010). Further we are able to direct priority areas to be concentrated on to balance both the needs of an expanding human population including poverty eradication and food needs as documented through the United Nations Millennium Development Goals (http://www.un.org/millenniumgoals/ , accessed July, 2013) and the continued distribution of richness of plant species.
.
References
Arrhenius, O. (1921). Species and Area. Journal of Ecology, vol. 9, pp. 95-99.
Barthlott, W., Mutke, J., Rafiqpoor, D., Kier, G. and Kreft, H. (2005). Global Centers of Vascular Plant Diversity, Nova Acta Leopoldina NF, vol. 92, pp. 61–83.
Bhattarai, K. R. and Vetaas, O. R. (2003). Variation in plant species richness of different life forms along a subtropical elevation gradient in the Himalayas, east Nepal, Global Ecology and Biogeography, vol. 12, pp. 327-340.
Büche, D., Schraudolph, N. N. and Koumoutsakos, P. (2005). Accelerating Evolutionary Algorithms with Gaussian Process Function Models, IEEE Transactions of Systems, Man and Cybernetics – Part C: Applications and Reviews, vol. 35, pp. 183-194.
Furze, J. N., Zhu, Q., and Hill, J. (2013). The use of plant characterisation studies to substantiate national conservation and sustainability policies, Global Education Magazine, issue 5, pp. 68-75.
Furze, J., Hill, J., Zhu, Q. M. and Qiao, F. (2012), Algorithms for the Characterisation of Plant Strategy Patterns on a Global Scale, American Journal of Geographic information System, vol. 1, no. 3, pp. 72-99.
Furze, J. N., Zhu, Q., Qiao, F. and Hill, J. (2013a). Implementing stochastic distribution within the utopia plane of primary producers using a hybrid genetic algorithm, International Journal of Computer Applications in Technology, vol. 47, pp. 68-77. DOI: 10.1504/IJCAT.2013.054303
Furze, J. N., Zhu, Q., Qiao, F. and Hill, J. (2013b). Mathematical methods to quantify and characterise the primary elements of trophic systems, International Journal of Computer Applications in Technology, vol. 47, pp. 315-325. DOI: 10.1504/IJCAT.2013.055324
Furze, J. N., Zhu, Q. M., Qiao, F. and Hill, J. (2013c). Linking and implementation of fuzzy logic control to ordinate plant strategies, International Journal of Modelling, Identification and Control, vol. 19, pp. 333-342. DOI: 10.1504/IJMIC.2013.055651
Furze, J. N., Zhu, Q., Qiao, F. and Hill, J. (2013d). Functional enrichment of utopian distribution of plant life-forms. American Journal of Plant Sciences, (in press).
Grime, J. P., Hodgson, J. G. and Hunt, R. (1995). The abridged comparative plant ecology. Chapman & Hall.
Grime, J. P. and Pierce, S. (2012). The Evolutionary Strategies that Shape Ecosystems. Wiley-Blackwell, Chichester, UK.
Hawkins, B. A., Field, R., Cornell, H. V., Currie, D. J., Guégan, J. F., Kaufman, D. M., Kerr, J. T., Mittelbach, G. G., Oberdorff, T., O’Brien, E. M., Porter, E. E. and Turner, J. R. G. (2003). Energy, water, and broad-scale geographic patterns of species richness. Ecology, vol. 84, pp. 3105-3117.
Jenkins, C. N., Pimm, S. L. and Joppa, L. N. (2013). Global patterns of terrestrial vertebrate diversity and conservation, Proceedings of the National Academy of Science Early Edition, vol. 110, 1-9.
Kraft, N. J. B. and Ackerly, D. D. (2010). Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian Forest, Ecological Monographs, vol. 80, pp. 401-422, 2010.
Kreft, H. and Jetz, W. (2007). Global patterns and determinants of vascular plant diversity, Proceedings of the National Academy of Sciences, vol. 104, no. 14, pp. 5925-5930.
MacArthur, R. H. and Wilson, E. O. (1967). The theory of island biogeography. Princeton University Press.
New, M., Hulme, M. and Jones, P. (1999). Representing twentieth century space-time climate variability. Part I-Development of a 1961–90 mean monthly terrestrial climatology, Journal of Climate, vol. 12, pp.829–856.
Prusinkiewicz, P. and Lindenmayer, A. (1990). The algorithmic beauty of plants. Springer-Verlag.
Raunkier, C. (1934). The Life Forms of Plants and Statistical Plant Geography, Oxford University Press.
Salisbury, F. B. and Ross, C. W. (1992). Plant Physiology Fourth Edition, Wadsworth Publishing Company, California, USA.
Schölzel, C. and Friedrichs, P. (2008). Multivariate non normally distributed random variables in climate science – Introduction to the copular approach, Non linear Processes in Geophysics, vol. 15, 2008, pp. 761-772.
Simberloff, D. S. (1974). Equilibrium theory of island biogeography and ecology, Annual Review of Ecology. Evolution and Systematics, vol. 5, pp. 161-182.
Takagi, T. and Sugeno, M. (1985), Fuzzy identification of systems and its applications to modeling and control, IEEE Transactions on Systems, Man, and Cybernetics, vol. 15, no. 1, pp. 116–132.
Trauth, M. (2006). MATLAB® Recipes for Earth Sciences. Springer.
Wright, D. H. (1983). Species-energy theory: an extension of species energy theory, Oikos, vol. 41, pp. 496-506.
This article was published on 10th December: Human Rights Day, in Global Education Magazine.








